Perched atop the hill at Ocean Forest Ecolodge, the ceremonial lodge offers breathtaking panoramic views of the sparkling Pacific Ocean and the legendary Caño Island. Referred to as “the old hill… of painted designs,” this hilltop sanctuary honors both the sacred lineage of yagé and the warm, faith-driven spirit of its keepers.
Immersed in mother nature’s magnificent presence the ceremonial space is essentially a roof over an earthen floor, this according to protocols of the tradition of yagé, and where fires can be lit for burning incense. Bamboo cots and poles allow for tying hammocks, and offer space to accommodate over 20 participants. Surrounded by original rainforest teeming with life and lush ethnobotanical gardens where sacred entheogenic plants are cultivated, the lodge offers a setting that allows a timeless imprint favoring spiritual growth and inner renewal of body mind and soul.
The maloca features an earthen floor—essential for a ceremonial lodge. It offers grounding, provides a hearth for sacred fire, incense, and smudging, and plays a vital role in yagé ceremonies, naturally absorbing the purging that often accompanies the medicine.
This peaceful sanctuary offers space to unwind, take in stunning views, hike rainforest trails, and enjoy daily visits from abundant birds and wildlife.
Here at the ceremonial lodge, yagé is prepared always according to traditional standards, honoring the sacred “Designs Path.” To learn more, read “Preparing a Proper Brew” in Rainforest Medicine.
The grounds feature a thoughtfully designed composting toilet with an open-air vista, overlooking a verdant rainforest valley. And from the toilet, my oh my, what a view! The compost, once ready, goes to the fruit trees planted at the site, who really appreciate the nutrient rich fodder!
Here, bird and wildlife encounters are common with monkeys, toucans and white hawks seen on a regular basis. Many other birds can be seen here as well, such as daily scarlet macaws that fly past, colorful tanagers, cuckoo birds and different species of woodpeckers. Impressive wildlife encounters are common, such as with the daily visitation of white faced and howler monkeys, groups of coatimundi, kinkajou, tayra, tamandua, squirrels and the ocasional tapir will walk past. And where the cool nights of yagé can be experienced, in absolute tranquility, under the light of moon and stars.
It is a great joy and an honor to share this space during our Rainforest Medicine Council Gatherings as well as make it available for other visiting groups.
Dunduma, a spiritual plant, is believed to have originated from the heart of Shiu Amarun, the shimmering, multicolored and resonant fertility mother boa of the Earth, related Taita Casimiro Mamallacta Mamallacta, Kichwa traditional elder from the outskirts of the jungle town of Archidona, in Ecuadors upper Amazonian Napo Province.
According to legend, Shiu Amarun was slain by Atacapie, the seven-headed boa of chaos and disintegration, with each head pulling in a different direction. Atacapie symbolizes chaos and confusion, much like the disjointed nature of current governmental institutions. In contrast, Shiu Amarun embodied harmony and fertility, celebrated in many legends for representing the purity and balance of Mother Earth. After Shiu Amarun’s demise, seven plants sprouted from her heart, ensuring that her legacy and energy would endure. You can read the complete myth in my book Rainforest Medicine ~ Preserving Indigenous Science and Biological Diversity in the Upper Amazon, page page 70, “A Kichwa Legend of the Creation of the First Ingandu (Ayahuasca Vine).”
These seven plants are:
Ayahuasca (Banisteriposis caapi)
Amiruka Panga (Psychotria viridis)
Dunduma (Cyperus sp.)
Uchu (a variety of red hot chili pepper ~ Capsicum sp.)
Iru (a variety of sugarcane ~ Saccharum sp.)
Paju Jinjibri (a small spicy variety of ginger ~ Zinziber sp.)
Palanda (a small variety of banana ~ Musa paradisiaca var.)
Dunduma sprout
Dunduma holds profound significance in the Amazonian tradition, with many of its uses passed down through generations of elders. The plant is magical and revered among the natives of the upper Amazon. Its Kichwa name originates from the Upper Napo Province, particularly around the town of Archidona. The Siekopai call it “Yiyó, Nuní,” meaning “Celestial tuber of beads.” In their cosmology, the bead represents the union of opposites, with its empty hole symbolizing the truth that matter is made effective by spirit. Here are some of its commonly known uses:
Protective Spray: The root is mashed into floral water, cologne, or cane moonshine to create a protective spray. This spray is applied to individuals when they feel dizzy, experience blood pressure issues, or need protection during travel or hunting. It is also used for good luck in meetings and to revive someone who has fainted.
Clarity Ritual: The roots can be grated and soaked in hot water, which is then cooled and strained. This water is consumed like Kava during dawn rituals, helping participants discuss dreams, plan for the day, share legends, and address life issues. Dunduma is believed to grant clarity and lucidity, helping one understand problems and find solutions.
Healing Tonic: Dunduma soothes upset stomachs, colic, menstrual cramps, dizziness, anxiety, and general unease. It is safe for children, with a small dose for babies and a half-cup for older kids.
Enhanced Potency: The water’s potency is enhanced through chanting, praying, singing, or leaving it out under moonlight. Elders with knowledge of specific invocations can further amplify its qualities.
Pain Relief: Dunduma is also used to relieve body pain, particularly muscle aches and cramps. The water or essential oil can be applied to painful areas, and it is beneficial for heart pain when prayed over and consumed. A balm can be made by mixing the extract with lard or fat for massaging stiff or aching body parts.
Support for Children: Dunduma is believed to help children overcome laziness caused by anemia and purify the blood, strengthening the heart.
Antiparasitic: Regular consumption of Dunduma helps rid the body of parasites due to its anti-parasitic properties.
Fungal balls on a mature Dunduma
The name “Nuní” refers to a collection of celestial plants in the genus Cyperus, all with aromatic tubers, which are ancestrally cultivated heirlooms believed to contain the holy spirit of divine immortals.
In Colombia, some groups refer to Dunduma as “Trueno Chunduru,” the sacred seed of lightning and thunder, emphasizing its awakening properties. Like thunder clears stagnation and brings freshness, Dunduma cleanses one’s aura and energetic space, restoring clarity.
Wati Nuni: A Sacred Plant of Spiritual Protection and Healing
Watí’nuní
Description and Symbiotic Nature
Wati Nuni, a Cyperus species and a variety of Dunduma, is distinguished by its thin, tube-like stalks with a spurts of lanceolate leaves protruding out from the tip of its stalks. This plant forms a symbiotic relationship with native fungal species, which appear as white balls of fungus at the center of the leaf cluster. These fungus balls were traditionally used by shamans in spiritual healing practices.
Spiritual Significance
Wati Nuni is revered for its mystical properties, known for its ability to unite all the chakras and align one’s spiritual essence. This sacred plant is believed to protect the home and ward off evil spirits. Its water, made from chopped roots soaked in water, is similarly effective in repelling negative energies and for a wide array of ailments, anxieties and distrubances. Being a gift from celestial beings, its purpose is to upgrade harmony and protection.
Cultural Legacy and Meaning
The name “Wati Nuni” translates as “Wati” (spirits of all kinds) and “Nuni” (sacred heavenly seed). These plants are considered gifts from divine immortals, brought back by spiritual masters during visions. Wati Nuni is part of the legacy left by yagé drinkers, who passed down these sacred plants to aid ordinary people in their spiritual cultivation, protection, and overall well-being. This is so being that inside the very tissues of his plant is enshrined the holy spirit of the divine immortals.
Ceremonial Use
Wati Nuni, along with Dunduma, plays a significant role in ceremonial settings, particularly after consuming ayahuasca. It is used to wash the mouth, a practice believed to calm and protect the participant. The water in which the seeds are soaked can also be splashed around the ceremonial area using a ti leaf sprout, protecting the space from malicious spirits. This aromatic water is used as a protective spray, and when blown on the temples or vrown its calms, and opens sacred space for meditation and communoin wiht the always new and fresh divine immortals. Cultivating Wati Nuni is regarded as a spiritual resource and a blessing, with the plant opening pathways for blessings to unfold based on the sincerity and virtue of those who grow and use it.
Preparation and Rituals
To prepare Wati Nuni for use, the roots are sliced into small pieces, soaked in water, and prayed over. Although the plant has inherent power, its potency can be significantly enhanced through intentional rituals such as leaving the water under moonlight, chanting, and praying. Specific prayers must be directed for the plant to perform its intended work, whether for protection, healing, or aiding someone in need. Wati Nuni is also used to treat spirit possession, help children with autism, and ease anxiety and fear.
Benefits for Children
Wati Nuni is believed to instill natural discernment between right and wrong in children, especially before they go to school. This quality helps them resist bad influences. The water is given to babies if they are frigtened or uneasy and is like chammonile, soothing and calming.
Boundary Protection
Water infused with the chopped tubers of both Dunduma and Wati Nuni can be applied to boundary lines during disputes, serving as a protective barrier against intrusion. However, this remedy should only be used in legitimate cases of boundary disputes, invoking the plants’ protective qualities to restore harmony.
Cultivation
The Siekopai people cultivate this plant near their homes in full sunlight, using soil mixed with ashes from the fire. When the leaves dry up, the tubers are harvested, cut from the roots, and placed in water to sprout before being replanted.
Conclusion Both Wati Nuni and Dunduma are versatile plants, capable of thriving in various climates but requiring full sun exposure. Excessive vegetation can lead to their demise, and over time, many culturally significant varieties have been lost. For example, there were once varieties used in rituals to make the deceased visible to divine immortals, guiding their souls to the celestial realms.
One fascinating aspect of Wati Nuni is its vast genetic diversity. When planted together, it can grow into small, medium, and large varieties, each serving unique purposes. The dwarf variety is known for breaking spells, the medium-sized variety is used to resolve social issues, and the tall variety helps those who feel disintegrated by aligning all aspects of their being.
These sacred plants serve as agents of the co-evolution of heaven and earth, opening celestial pathways for those who tend to them with sincerity and spiritual discipline.
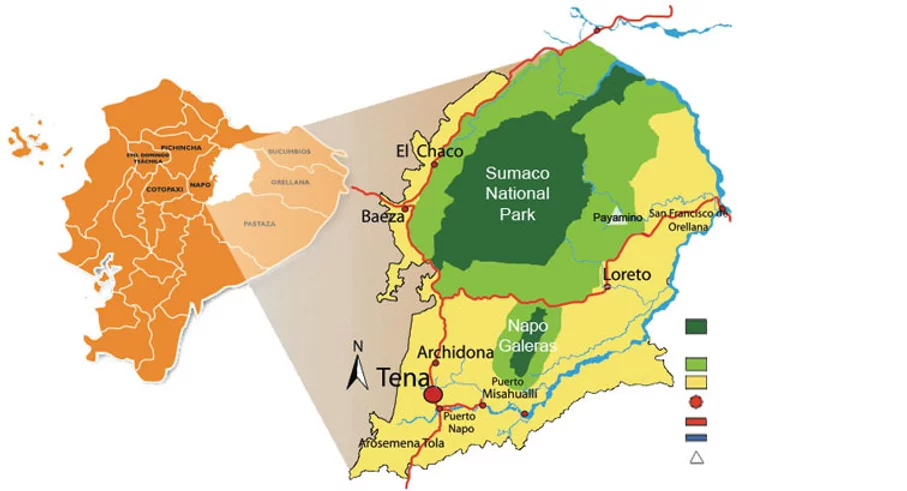
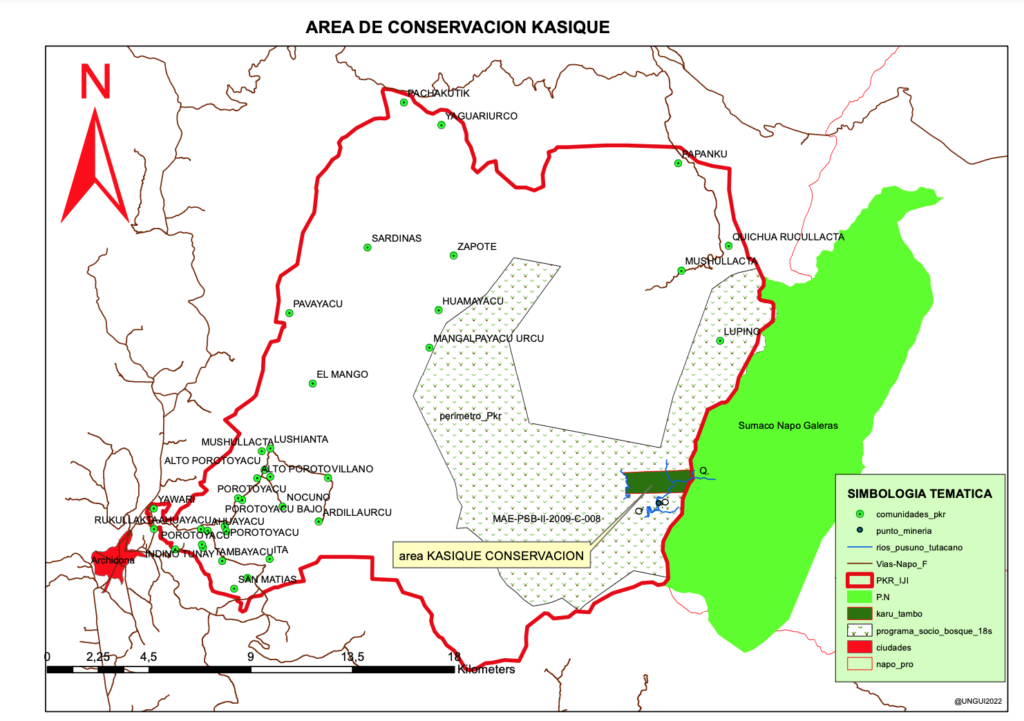
Conservation Adventures at the Epicenter of Botanical Diversity…,at the “Great Earthen End of the World Jaguar Mountain,” the isolated limestone massif ~ today known as: Cordillera Napo-Galeras, part of Sumaco Napo-Galeras National Park.
The mountains indigenous names translate the powerful vision regional peoples have for this sacred place, knowledge that may little be passed down to the younger generations, yet once not long ago was held as central belief among the people. Kichwa wisdom keepers relate the mountains ancestral name to be, Izhu Mangallpa Urku, “The Great Earthen End of the World Jaguar Mountain.” Among the Waorani, not the literal translation of Eygahueyaboga, which is “Mountain of the Eygahue bush,” an endemic bush, rather the deeper translation revealing the mountains far reaching meaning in accordance with oral tradition and Waorani mythology is, “People-who-have-swayed-from-original-instructions-devouring-Jaguar-mountain-of-the-most-powerful-revival-leaf.” Another name the mountain goes by is Unkiyabe, meaning “The-house-of-the-immortals-of-the-inner-earth-dimensions” or “The longhouse of the monakageiri,” these being a legion of short extremely high energy ancestor immortals who are progenitors of the Waorani people.
Mythological scene of the mountain and the “The Great Earthen End of the World Jaguar” emerging from the sacred mountain of Napo-Galeras to devour offending humanity. (Sketch by Thomas Wang – From the book Rainforest Medicine)
The High Hill ~ House Mountain ~ Cordillera Napo-Galeras
In the early 90’s I met Casimiro Mamallacta Mamallacta, a wholehearted Kichwa elder and traditional adept from the outskirts of Archidona. I wrote about Taita Casimiro and some of the experiences shared and learned from this happy camper in chapter 8, “Eyebrows of the Andes,” in my book Rainforest Medicine – Preserving Indigenous Science and Biological Diversity in the Upper Amazon. Between the years 1990-1994 we focused our efforts, in a way much like placing a magnifying glass in the sun to concentrate a beam of energy, into an inspired focus, capable of ensuring the ecological integrity of a place of great environmental and culture importance. Cordillera Napo-Galeras, not just an isolated limestone massif rising up from the lowland Amazon 80 kilometers east of the Andes. Not just a place rich in endemic species at the epicenter of botanical diversity. Yet as well a place enshrouded in profound ancestral belief among the region’s Indigenous peoples; most particularly among the community wisdom keepers, as a holy site with mythic connotations attesting to the veracity of the great deluge, while endorsing premonitions of yet another apocalypse.
Aromatic ShikaShika YakuA Slipper OrchidA Biodiversity WonderlandFossils of an ancient seabed
A Biological Diversity Wonderland Woven in a Mythic Fabric
Not just a place rich in endemic species, at the epicenter of botanical diversity, yet also, a place woven in a mythic fabric among the regional indigenous peoples. Cordillera Napo-Galeras, known as the “Great Earthen End of the World Jaguar Mountain,” where from within will emerge the “Great Earthen End of the World Jaguar,” the Izu Mangallpa, or Mundo Puma, that Runashimi ethnopoetics relates will devour offending humanity. Among the Siekopai the mountain is known as Jaicunti, the “High Hill,” within which abide the Jaicunti Pai, “Immortals of the High Hill,” the unfailing, ultra immaculate, fluffy clean medicine guides of the ceremonial plant medicine tradition of Yagé. Healers associate with these immortals to accomplish their effective spiritual cures. And to the Woarani, the mountain is known as Unkiyabe, “House Mountain,” a place where in times of old, Waorani families escaped the great deluge. Mengatue the Jaguar Shaman related that inside this mountain live immortals known as the Monakagaeiri. Elders relate them to be small in size and that they once lived among humanity benevolently teaching the early people how to live in the wilderness. The mountain is also known among the Waorani as Ewhengawehyabo, the “Humanity Devouring Jaguar-nip Mountain.” Waorani mythology relates that when heaven and earth split a celestial goddess brought the Waorani to the mountain, indicating to them the limits of their earthly territory. She informed them that when all fails, it will be from here where they are to request her intervention. She requested that from here nothing be taken. Heeding the warning, a mischievous viewer, took a branch from the tree where Jaguars of many sizes where lounging. When the plant at his community below sprouted, out of the plant came a giant jaguar that devoured half the village! When the plant was cut down, the giant jaguar swiftly loped back to the mountain, where it disappeared, inside the mountain itself.
The magnificent Pusuno River, at the foot of Napo-Galeras, the “River of Bubbles” where pilgrims come to purify themselves at the wee hours of dawn, before the hummingbird swims, and to drink Guayusa.
At the Epicenter of Botanical Diversity
When I first started hiking to Napo-Galeras it was an arduous several days trek from Archidona, and through literally knee deep mud! As the years passed new roads pushed deeper and deeper in the wilderness. One of the roads, the Hollin-Loreto road, we can sadly say has been an absolute ecological disaster, opening an exquisite wilderness area at the base of the Andes to a flood of human intervention. Piercing through a remote virgin valley between the mountains of Sumaco volcano and the cordillera Napo-Galeras, an area that soon botanists from the Missouri Botanical Gardens would determine to be the epicenter of botanical diversity.
As colonist settlers rushed into the area to claim lands the rainforest began shrinking in the wake. The original protection category granted to Sumaco and Napo Galeras region in 1987 shrank in size by three quarters when the two mountains were declared a National Park in 1994 losing millions of acres of rainforest to the impact of human settlement complexities. I might add, the original protection status came in 1987 when USAID gave the funds for the bridges to built the Hollin-Loreto road. Since then small towns like Huaticocha have now burst in size with the swift passing of time. And It was here that in the year 1994, from within the limits of the Napo-Galeras National Park, that I oversaw a process that relocated six colonist settler families. It was promised to them that if we could not improve their situation, then we would do nothing. Two large farms near Huaticocha were purchased and divided into lots for these settlers, and we won their hearts over forever! That was an adventure all right!
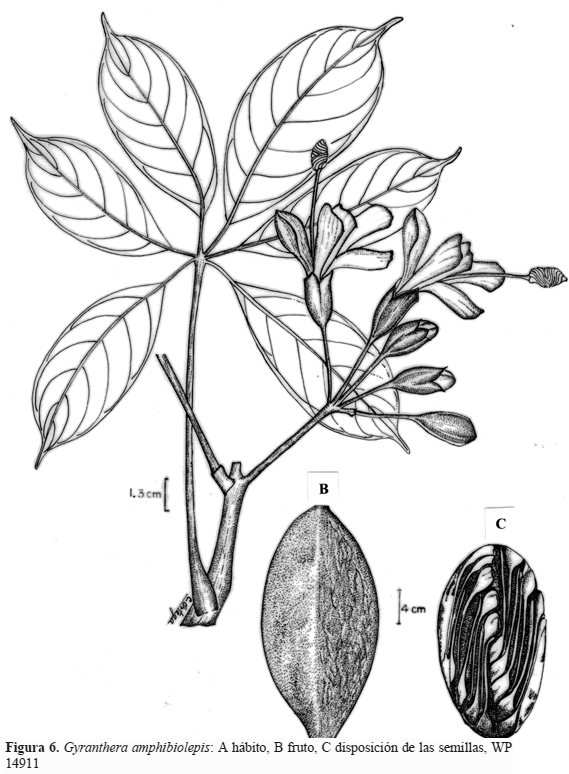
The new roads made possible the pillage of the old trees, and giant trees of many kinds would be seen on trucks being hauled to the mills. One among these, is Kamotoa. A towering canopy tree reaching heights up to 40 meters (131 feet) as yet unnamed to science. A locally endemic tree found only within a narrow band along the eastern base of the Andes, on limestone subsoils; today nearly extinct due to unregulated logging. Amazingly so this tree has only recently been described in 2002, named by Ecuadorian dendrologist Walter Palacios as Gyranthera amphibiolepis; pertaining to the Malvaceae botanical family.
Demarcation and Protection of Napo-Galeras: 1990-1994
Cordillera Napo-Galeras
Our work to demarcate and protect Napo-Galeras mountain, as if invoked into existence by the holy spirit of the mountain itself, had been occurring simultaneously, but not as a part of, an initiative being spearheaded by the GTZ that was pushing for higher protection status for the region. This organization with funding from the German government was pressuring for higher conservation measures for the Sumaco volcano. This alongside multimillion dollar grants to the park service for the region to receive National Park status, Ecuador’s highest conservation agreement. We heard from contacts in the park service that there were no maps for Napo-Galeras and it ran the risk of being left out of the soon to be declaration to National Park status. It was clear Napo-Galeras was being omitted from this plan. The need to present an irrefutably convincing technical report pitching Napo-Galeras be included into the upgrade to National Park status, became clearly evident.
We had already accomplished the maps showing the actual ground level limits of the Kichwa communities surrounding the cordillera Napo-Galeras. Showing what could be declared as a higher protection status without being rejected due to the excuse of human intervention. Our mapping project revealed that the majority of the cordillera Galeras, 14,687 hectares remained free of human claims, as well as 3,700 hectares of an important low land sector within the Tropical Wet Forest bioregion. We had already worked to relocate six colonist settlers living within the reserves northern boundaries, leaving the entire limits of the park surrounded only by Kichwa speaking comunas. Who, I might add, are also colonist being they were able to access the region due to the new Hollin-Loreto road. Some Kichwa of the area do have much older claims on the land though as the region north of Galeras was a hunting territory in the past, and a travel route to the old Kichwa Village of Avila Viejo on the flanks of Sumaco volcano. The region east though where many Kichwa communities are now located was once part of the Waorani territory. This is clear with the history of the Kichwa Village of San Vicente del Río Bueno, closer to the Napo-River, where Vicente Legisamo was speared to death by the Waorani in the mid 70’s. The story was told to me by Vicente’s brother Eduardo, the first colonist to settle in the area. He married a Kichwa women and brought Kichwa speakers from Tena to populate the region.
Examples of non sustainable land use that is sadly the status quo available to locals in the area. Naranjilla cultivation relies on heavily being sprayed which depletes the virgins soils. The people end up with next to nothing despite how hard they work.
After overseeing the process to relocate the six colonist families, we began another phase of the project to map and explore the area on Galeras eastern slopes. The first time we went in to visit the area known as Santa Rosa de Arapino, we got good and thrown out by the local comuneros. (Comuneros are the people who comprise the local Kichwa speaking communities today found in the region.) They had heard what was happening with the relocation of the colonist settlers at mountain’s northern boundary in the Pucuno River gorge and thought we were there to do the same to them. They escorted us to their community boundary, on a long hike through the mud in silence. Just as we were to depart, I said, “We will gladly leave, just keep in mind though, that were the only ones who will ever come here in the name of the holy mountain, in the name of the end of the world jaguar itself, and were only here to help you protect this place.” It wasn’t long before they sent message through the town mayor asking us to return, and we were back on the scene. It was clear that the only way to have the surrounding comuneros support the initiative was to inspire them to take an active role in protecting what was still left unclaimed. And we were able to gain their support when they saw we were there to legitimize their community boundaries and share ideas that they resonated with.
Village of Mushullacta “New Village”
Soon we returned and the community meetings began. We explained the National Park must be seen as a nest, out from which many good things come, everything inside the nest though must be protected, it must be left just as it is now, in its original and virgin state. The comunas had claimed exquisite lands and it was wild to see such a place and know that it was the beginning of a major ecological transition about to occur. Large families with dismal economies needing to see how to make it off the land, in the heart of biological diversity, with sadly only unsustainable and ecological degrading options at hand. We spoke of ecotourism potential and of the benefits of living next to a National Park and in light of this they will eventually receive support from government programs for their communities in parks buffer zones. Maybe not yet but as time progresses and these areas gain more attention, they will receive support for community projects. They asked us to analyze the economics of the logging going on, and it was deduced quickly that they are being highly taken advantage of. Colombian loggers would give them a chainsaw in exchange for five large trees, after which the chain saw would break and they would have no trees and no saw.
My field sketch of C. cateniformis
Massive Chunchu trees (Cedrelinga cateniformis), considered among the Siekopai as a sacred tree called Monsë, “Day Tree,” being butchered for chump change. Legend relates that the Monsë tree grows at the center of the sacred hoop of life, among the Siekopai the bark is used as an emetic to cleanse oneself for the ceremonial imbibition of the ancestral rainforest plant medicine of Yagé. We recommended they halt immediately all logging and sell the trees not once, but a thousand times via ecological tourism, that also rescues the culture. We held Permaculture courses and spoke of soil regeneration and sustainable agriculture. We shared knowledge of medicinal plants and held ethnobotanical gatherings aimed at raising the common body of botanical knowledge. We spoke about alternative economies and about improving their cacao and making products; that they must focus on opening markets for their crafts. Essentially we spoke about the need to become more self-sufficient and less reliant on the outside world. As it will be this that ultimately brings them the most security.
And we spoke of the legends and cultural implications for Galeras and that we sincerely hoped they would see it as a holy gift from our creator to be stewards of this sacred place. We prayed together and drank chicha and celebrated too. They had never had anyone come around and give this kind of attention; right as these fledgling communities were taking form.
Wairachina Sacha ~ The Rainforest of the Purifying Winds
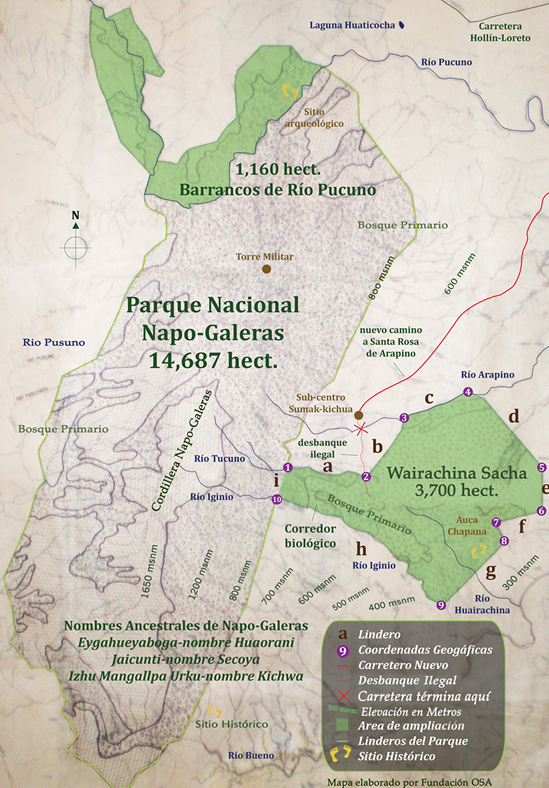
Map of Galeras and Wairachina Sacha
This process also revealed a very important reality. This being that a significant portion of the eastern lowland sector, that of the Wairachina Yaku watershed had still been unclaimed. An area we came to call Wairachina Sacha, “The Rainforest of the Purifying Winds.”
After further study it was revealed that behind Wairachina Sacha were land claims that would separate the possibility of an undisturbed corridor from the lowlands at 300 meters above sea level to the cordilleras high ridge at 1650 meters. It was preserving this intact gradient that had my passion glued as to what to do next.
To resolve this we managed to secure a biological corridor, as seen on this here map, by actually paying out one of the community members and having the community allocate the family a new lot in a different part of their community land claim. All this work came after many community meetings and socialization with the surrounding villagers. At the beginning it was rough. Thanks to grandfather Cesar Mamallacta, a man with a heart of pure gold, who accompanied us. Grandfather Cesar spoke in the native language of Runashimita; deeply from his heart and highly in our favor, reiterating time and time again that we come in the name of the mountain, in the name of the birds, animals and trees. That we come with an idea that will not perish, for the vision we share is one of goodness and beauty.
Accused of having trapped the World Puma
in a net; for having brought it to a zoo in Quito
At one such meetings, a man came out slightly drunk, and began accusing me. He said that I had caught the Mundo Puma in a net, another name for the End of the World Jaguar that is believed to reside inside the mountain, and that I had brought with a helicopter to a zoo in Quito. Eventually I was able to respond when it was my turn to speak. I replied saying that if that were the case I’d have no reason to be here now bothering you. I’m here to inspire you to protect the holy mountain of Galeras to ensure the Mundo Puma does not come out and eat us all! Everyone cheered in joy and more and more came to listen to the ideas of the crazy gringos, on fire with passion for the rainforest! Another villager hugged me crying in joy. He said that he agreed deeply with everything we spoke of, and it was so refreshing to hear what he always knew to be correct now being spoke of for the first time among the entire village.
Amazingly so, with a shoestring budget and rubber boots full of passion, our work helped consolidate the boundaries of what could now be protected. The above work took, I might add, the greater part of 4 years, this being between 1990-1994. Sure enough the work was strong enough to have the government include Napo-Galeras into the National Park status, and the new park came to be known as Sumaco Napo-Galeras National Park. This was epic indeed! Also amazing to see was how our initiative, called Demarcation and Protection of Napo-Galeras Mountain was able to inspire all the communities surrounding Napo-Galeras mountain and Wairachina Sacha to participate in the conservation and protection of the region to ensure others would not claim the land inside the protected areas and hopefully live softly in this precious rainforest region they now find themselves in.
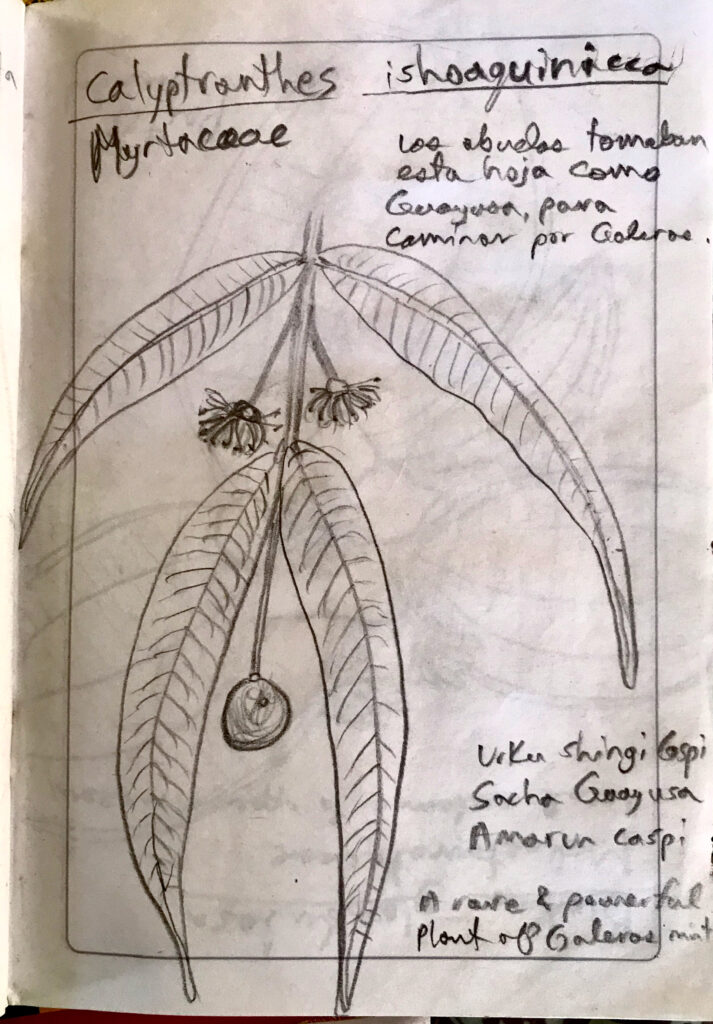
Some botanical sketches I made while in the field. L-R: “The strong bark of the mountains” ~ Calyptranthesishoaquinicca, 2. Blakea glandulosa a local endemic, and a new species yet named in the genus Salpingia a lithophyte from Galeras summit.
Getting Wairachina Sacha Back on the Conservation Map: 2010 Initiative
And this effort lasted 16 years! Until the park service went to go clear the limits of the National Park in 2010. Only then did all the comuneros see that the lowland sector of Wairacina Sacha, was not actually “inside” the park. Wairachina Sacha is outside the legal limits of the National Park, yet within the limits of the first conservation status placed onto the region, that of a Patrimonio Forestal. Among the most priceless regions of biodiversity in Ecuador’s upper Amazon was being dropped like a rock into the river. And then wala we show up again! And we were there on a big fishing mission indeed! The only Tropical Wet Forest remaining that could be included in a National Park in Ecuador’s Amazonian region. This is so due to the fact the Ecuador’s Amazonian protected areas are all below 250 meters elevation and above 800 meters elevation. The critical region of biodiversity, the Tropical Wet Forest, that is the overlapping edge between the lowland Amazonian Tropical Moist Forest and the rainforest of the Andean slopes, an area rich in endemic species, the origin location of cacao, not being represented in the SNAP – Ecuador’s System of National Protected Areas. Seeing all this led to a whole new conservation adventure!
A new species of Aiphanes. Dr. Cerón identified one of my collections as a new species of palm from Galeras summit.
In light of this, in 2010 an inspiration arose to engaged in yet another Rainforest conservation adventure. With the encouragement of my old friend Benjamin Mamallacta I was inspired to return. The mission now was to reevaluate the situation; initiate a process to secure greater protection status for this priceless biodiversity region area east of the Napo-Galeras massif. This time though we discovered an illegal road about to pierce right through where in 1994 we successfully negotiated the biological corridor. Fortunately the tractor sank so deep into the mud they were obliged to halt, then we used legal resources to stop it once and for all. It just happened to be the UNESCO International Year of Biodiversity as well and we wrote letters to UNESCO, who actually did reply, requesting the government consider our petition. The new park service maps no longer reflected the area to have the original conservation status from 1987, that of forest patrimony, that under Ecuador’s new constitution now has a much higher protection status than when we started in the early 90’s. Via petition signing, an information campaign and many meetings with local government representatives. Complete with ground level commissions that brought representative from the park service to see the region first hand. Being the road was illegal, it was fortunately swiftly stopped! Wairachina Sacha as a patrimonio forestal was put back on the new conservation maps under the category of buffer zone and the need to protect this place was again corroborated on a governmental level.
Landslide on the Hollín-Loreto, new roads into Kichwa villages, Illegal road into Wairachina Sacha and Sumaco Biosphere map.
Amazingly so and much to my surprise the communities had still not let anyone claim the area of Wairachina Sacha. They had remembered us from 1994 with fondness and we lifted up yet again another round of community meetings focused on re-socializing the need to continue protecting this region among the local community members and neighboring comunas. It was agreed by the surrounding communities that this they would continue to do! There’s a lot more I can write about this process, yet for the meantime, I will leave you here only with the end result, that thanks to great spirit, and rubber boots full of passion, was successful! Self-funded from savings from our family operated rainforest and ocean ecological tourism and yoga center in Costa Rica and small donations from friends, we were able to accomplish, in my humble option, a whole lot!
As part of the multidisciplinary task at hand, a botanical expedition was organized with Dr. Carlos Cerón, curator of the Alfredo Palacios herbarium at Ecuador’s Central University. On this trip it was discovered that this area east of Galeras is the evolutionary center of the Malvaceae family, this being due to a disproportionate amount of plants in the Malvaceae family found. Including among these cacao and many cacao relatives. Do check out my previous blog entry about this fascinating topic, Cacao The World Tree and Her Planetary Mission.
Cacao (Theobroma cacao)Cacao artifact Mayo-ChinchipeBiodiversity displayMayo-Chinchipe Cacao vessel (3000 BC)Eyebrows of the AndesThe megabiologicaly diverse Tropical Wat Forest, a narrow band between 400-600 meters elevation, is considered to be the origin location of Cacao. In this region, 5000 year old Mayo-Chinchipe Cacao artifactshave been found, at the La Florida archeological site. Villagers here hold annual cacao festivals a fruit emblematic of the region. See my article: Cacao ~ The World Tree and Her Planetary Mission.
Then on the way out at the town of Huaticocha the following occurred, leaving on me an indelible memory. We were limited on gas, so I asked at a local tienda (small food shop) if they might have a few gallons they could sell us. A man sitting there looked at me attentively and suddenly called my name, “Jonathon!” It was one of the settlers that I had relocated in 1994. He remember me and with great joy and he siphoned the gas from his truck for us to use. He pointed at the ridge line of tree in the not so far distance, the Napo-Galeras National Park, and said, “See that line of trees? That is there because of you!”
Jungle hammock on the Osa, Costa Rica
He went on proclaiming, that I was the first and yet only one that has ever done anything important in their town. He handed me his car keys and said that any time I needed his truck or a place to stay, that everything he owned was faithfully at my disposition! He said that he had gone one to become an agronomist inspired by the talks we gave, and he mentioned the other settler families we had relocated from the deep wilderness to the two large properties we were able to purchase and divide among them near the town, had all devoted themselves to upholding the permaculture principles we had left there as a teaching. I thanked him deeply, and that it was he who has given me the biggest gift. To see that the efforts we gave, showed results, and over so many years! And I was able to return to my hammock in Costa Rica on the Osa Peninsula, on the banks of mighty Pacific Ocean, to the splendid Ocean Forest Ecolodge, with a robust cheek to cheek smile on my contented face.
The Mamallacta Family Biological Reserve : 2021
Mama Margarita Alvarado and Taita Casimiro Mamallacta Mamallacta (photo by Benjamin Mamallacta)
In August of 2021 I returned with a group of friends to visit the mighty Pusuno River, the river of bubbles, whose purity and beauty has so deeply on me left her impression. This time working with Benjamin Mamallacta we began a program to protect grandfather Casimiro Mamallacta family Purina Tambu, back to where it all began! Purina Tambu literally translates from Runshimi, the Kichwa peoples language, as “a place that is walked to,” yet the term means so much more. “A place that is walked to,” is a place far from the bustle of human bedham, a place where the the quietud of nature permates. It is much more than this, it is where the ancestral ways can be upheld, this relates to dieting with the sacred teacher plants in order to learn and to practice the art of concentration. We can deduce then, that the Purina Tambu, is the legitimate ancestral university, where the individual can receive teaching straight from source through plant allies and mother nature. And like this gain strength and wisdom to live the each and every day. Here on the western slopes of Napo-Galeras, 1000 acres of wilderness have been delineated, where the Mamallacta family are focusing their conservation efforts. In late 2021 the boundary lines of the reserve were cleared and we are just about finished fixing up our original base camp, the house we built in 2007. This was made possible thanks to a small grant we applied for and received from The Global Green Grants Fund. I will write a seperate blog to share more about this initiative, for now though, allow me to conclude with the follow.
Ammonite fossils in creekYaku Yutzu (Calliandra sp.)Napo-Galeras and communities on western sideKasike Mamallacta Purina Tambu
Napo-Galeras ~ Woven in a Rich Mythological Fabric
The mythologies associated with this mountain are profound, running deep in the psyche of the people. There are accounts of Cayu Runa (lightning man) whose testicles at Galeras are transformed to stone, of mythic dimensional portals that yachac (wisdom keepers) of old would use to transport themselves to festivals in the high Andes. Legends of the origin of plants that grew from the heart of Shiu Amarun, the glistening fertility boa of the Pachamama, after a brutal battle with Atacapie, the seven headed boa of chaos and havoc. And of the Jurujuri, ardent protectors of the wilderness, who sucked out the eyes of offensive hunters. And there is talk of the Ingaru Supai whose heart hangs in his hollow chest, from which was made the magical maianchi powder, from where grew the Pumallullu a mystical plant of the people used for jaguar transformation. And the stories continue, relating accounts of Waorani chasing even don Casimiro off in his youth, and as far as the western slopes of the mountain, at the Río Pusuno. Casimiro chuckled, feeling it was just their way of saying hi. And there are the legends of the hero twins, Kuilluru and Ducero, and how they passified the powers of nature to allow humanity to prosper before ascending to become the dawn and dusk star that is Venus. And it was they who trapped the Mundo Puma in a cave at the mountains summit, informing the people that a day would come, in order to reestablish planetary harmony, when it would escape; this time to devour offending humanity. These are just some of countless legends related to the mountain of Napo-Galeras, like the lines emanating outwards on the ammonite fossils there found, remnants of a millions of years ago seabed that Amazonia once was.
Huaticocha Lake
I will leave you with one more insight, the legend of the Lake of Huaticocha. The town of Huaticocha receives its name from a lake near the town, the Lake of Huaticocha. This small lake is an anomaly being that it seems out of place, in the middle of a relatively steep forest valley and it has a powerful legend, one now few remember. Kichwa elders relate that the following phenomena there occurred and not too long back. An indigenous family of the name Siquihua who lived there were being harassed by colonist to fulfill orders of pita fiber. On one such order they were not able to complete the timeline, worried of the reproach they performed an ancestral rite, a ceremony with the sacred plant medicine of ayahuasca. They made a new clay pot and drank copious chicha (a staple gruel made from tubers or gran) as if in a festive celebration, so much that they began to vomit which they did into the newly made clay pot. That night they drank their ceremonial entheogenic ayahuasca brew. During the ceremonial trance the shaman family leader spilled the vomit onto the ground around their home. Where the vomit touched the earth, a lake formed, and the entire family sunk into the lake, complete with their house, their garden and all their pets, where they to this day live in an alternate dimension. Locals relate that this lake is indeed enchanted, being that when people attempt to come close the sky darkens and it begins to rain.
The legend of Huaticocha lake is about seeking refuge. Refuge from the impact of colonialism. Much in the same way, as the spirit of the mountain has invoked into action the very own protection of the mountains ecological integrity. Seeing to it that this holy place receive Ecuador’s highest legal protection category. Allowing for the refuge of countless species and spirits, that claim this mountain wilderness as home. The conservation of Napo-Galeras, we might deduce, in a way not so evident to see, is staving off the apocalypse itself! Holding back the emergence of the “Great Earthen End of the World Jaguar,” that is believed will emerge on judgment day to devour offending humanity. Or maybe it has already escaped, as some elders believe. This in light of all the havoc and destruction prevalent in the world today. Leaving us not much else to contemplate outside the phenomenon of understanding the universal subtle law, and aligning to this. So that at the very least, we may not be lumped into the general category of “offensive humanity.” And the faith of course, is that it’s never too late to shape up, so do please keep in mind…the myth clearly states, that it is the “offending” humanity, who the Mundo Puma will devour!
Dedicated to Miguel Payaguaje and his extended family (including his father Delfín and his grandfather Fernando), as well as all the vine gardeners responsible for stewarding these sacred plants through time.
Miguel Payaguaje making cuttings of Wai Yagé to plant in a garden with pineapples, Sucumbíos Province, Ecuador, 2017. (Photos T. Baldwin)
Introduction
This essay will present the origins, evolution, and human co-history of the Malpighiaceae family of ethnomedically significant vines in order to shed light on the confusion and controversy surrounding these important taxa. The intention is to draw attention to the importance of preservation and support of the indigenous cultures that continue to have deep connections with these sacred plants before this knowledge is gone. Images created with the confocal microscope of these species are featured in the website Microcosms: A Homage to Sacred Plants of the Americas.
Originof the Malpighiaceae Family
South America separated from Africa around 100 mya. The Malpighiaceae family of plants originated in South America approximately 70 mya. The family has roughly 77 genera containing approximately 1300 species, ranging in form from vine, tree, or shrub. 88% of the genera are from the new world with 22% escaping to the old world. Quite a feat considering South America and Africa had been split apart for ~30 million years before this family was born. (See Davis et al and Davis and Anderson)
Ecology and Fruit
Many of the vines in this family function ecologically as pioneer and accumulation species that help promote soil fertility by rapidly colonizing open sunny areas and degraded sites. They climb to the tops of tall trees in search of sunlight. Eventually, they can become so heavy that the host tree collapses under the weight, thereby creating gaps in the forest canopy. When a vine reaches the treetop, it is perfectly located for reproduction. Once the flowers are pollinated, the resulting fruits, when dry, are shaken by wind and released, sending them helicoptering down to the forest floor below, where some of them will find new fertile ground to germinate. The fruits of Banisteriopsis and Tetrapterys species are samaras, which resemble maple seeds with a wing-like appendage that cause them to whirl as they fall so winds can distance them from the mother vine. Fruits with wings have evolved and have been lost over time several times in this family. Other genera like Diplopterys and Callaeum have some species whose fruits are not samaras, but rather reduced winglets with air pockets that help them float with currents downriver. Herbarium collections from the upper Amazon basin demonstrate a pattern of locations for these genera: they flow laterally following the river courses from west to east. The more Banisterioid species with winged-fruit (for example Diplopterys longialata syn. B. rusbyana) have a vertical distribution running north-south along the eastern Andes. This distribution could be explained partially by the prevailing wind patterns pushing the wind-dispersed species up against the mountains and then riding the escalator of Andean uplift. Interestingly, only family members with winged, wind-dispersed fruits such as the genus Tetrapterys are found in both the new and old worlds. How might this have happened?
Sprouting somaran seed of Diplopterys longialata var huambisa, photo by Julio Vicencio at Jambi Mayu, Ecuador.Tetrapterys mucronata (Photographs by N. Logan with permission from the NYBG.), 1a&b – Bahia, Brazil (Amorim, Forzza, and Sant’Ana 3047), 2 – Rio Negro, Venezuela (Davidse 27817)
New World to Old World Migrations
During the Tertiary period, Tetrapterys and a few other related genera of the Malpighiaceae migrated north into North America and then moved east across a northern tropical corridor until reaching Europe. As temperatures cooled, the migrating Malpighiaceae moved south to central Africa where they can be found today. Other disjunct populations may have migrated later as long-distance dispersal events, but it is currently unknown how or when these most unlikely events would have occurred. While in the New World, oil-gathering bees have had a long-standing relationship with the Malpighiaceae. The disjunct populations of organisms in the Old World quickly morphed to adjust to the locally available types of bees. This demonstrates the genetic variation and morphological fluidity of this family and its ability to rapidly adapt to new circumstances. An example of this can be seen in the glands on the flowers of these species shifting from solely producing oil, to a mixed function strategy by which some glands produce oil, while others produce sugar. This has allowed the attraction of multiple potential pollinators to help ensure successful reproduction.
Early Encounters with Plant Synergies
Evidence shows early human inhabitants of South America combined tryptamine-rich seeds of the tree (Anadenanthera peregrina) with the beta-carboline-containing vine (Banisteriopsis caapi) to achieve synergistic effects analogous to the modern concept of ayahuasca (see Torres 2018). This could have happened very early in the migration process, because humans entering northern South America through the Orinoco river valley would have traversed large savannahs full of Anadenanthera peregrina, while B. caapi vines could be found growing at the edge of the zone where the savannah overlaps with the forest. It is reasonable to assume that in the search for food, both plants would have been an attractive source of nutrition for humans: water (vine) and calories (legume endosperm). Presently, some people of the Orinoco are known to chew raw caapi stems. In fact, the Piaroa, an Orinoco-based culture, still insist upon the use of Anadenanthera combined with B. caapi for the strongest effects (Rodd 2002). Cultural and ecological context would shape the recipe and method of consumption (Rodd 2008). Over time, methods have evolved, leading to the development of preparations that produce various states of consciousness associated with each respective context.
Ultimately, the use of B. caapi combined with more than one hundred potential admixture plants, became common across the eastern Andes from Bolivia, north to Colombia, and Venezuela, following the Amazon and its tributaries eastward across much of north and central Brazil. “Caapi” or “Cabi” are two of the more common names for referring to related vines across most of northern South America. B. caapi is considered by many groups of these regions to be a kind of driver of ecological ingenuity. It is the fundamental master medicinal plant teacher around which all other plants revolve.
Considering that the bulk of the vine speciation likely has occurred as a result of Andean mountain-building giving rise to novel microclimates, gravity and the physical barrier created by tall mountains would impose directionality to the spread of these species. Consequently, most migrate downstream from west to east. In Brazil, there are no verifiable occurrences of endemic Banisteriopsis relatives associated with traditional human use. For these reasons, the epicenter of the human-yagé-complex relationship is likely to be found in the upper Amazon basin at the source of the watershed.
The West Meets Caapi
For 15 years (1849 – 1864) English botanist Richard Spruce traveled upriver from the mouth of the Amazon beginning in Para, Brazil. He arrived at the upper Amazon basin (year ~1852) and traveled up the Río Negro as far north as the Orinoco (where he witnessed the chewing of caapi stems) and penetrated deep into the Río Vaupés observing and documenting the indigenous cultures (Guahibo, Tukano, and others) he encountered. From a garden plot, he collected and described Banisteria caapi (the genus later re-named Banisteriopsis) in addition to the cultural uses, rituals, etc. surrounding it. He collected a distinct variation of B. caapi with elongated swollen nodes. Spruce’s description of ayahuasca’s effects is different from modern descriptions in terms of onset and duration. The brew he encountered consisted (as far as we know) of caapi mixed with the slender lateral roots of a plant he believed was from the Apocynaceae (Iboga family), called caapi-pinima (painted caapi), a reference to the red-colored veins running through its leaves. Spruce first identified this admixture plant as a species of the genus Haemadictyon, later reclassified as Prestonia amazonica. While Spruce insisted the plant was of Aponcynaceous origin, the leaves he describes (sans red pigment) have an uncanny resemblance to Diplopterys cabrerana leaves, whose slender lateral roots combined with B. caapi stems are consumed as a decoction by the Witoto people. Not long after Spruce’s departure from South America, the rubber boom would begin, ushering in a new era of ecological degradation, cultural exploitation, and more western explorers looking for new natural resources to feed industry.
Schultes’ Enigma
Approximately 80 years after Spruce, economic botanist Richard E. Schultes was sent to the Río Vaupés to study arrow-poisons and collect new sources of rubber for industries supporting the war effort. He documented many important plants and ultimately inspired many people to become interested in ethno(sic)botany and help with the Amazonian cause. He documented and collected 2 different yellow-flowered, Malpighiaceous vines belonging to the caapi complex. Both B. martiniana (Isotype of Banisteriopsis martiniana (A.Juss.) Cuatrec. var. subenervia Cuatrec.) and the type collection of B. cabrerana (syn. D. cabrerana) were procured from tributaries of the Río Vaupés. Schultes made an effort to identify the painted caapi (Prestonia amazonica). Ultimately, he eliminated Prestonia amazonica (or any Apocynaceae) from the list of possible candidates of important admixtures to the caapi brew. However, he was unable to definitively ID Spruce’s painted caapi, ascribing this moniker to a species of Tetrapterys instead. He referenced at least 30 kinds of (known) named varieties of caapi in the NW Amazon. However, Schultes was baffled by the local indigenous people’s ability to identify different varieties of B. caapi consistently, even at a great distance without touching, smelling, or tasting the vine, whereas he was unable to distinguish them. This has been dubbed “Schultes’ Enigma”.
“This aspect of ethnobotanical studies certainly requires much more intensive and interdisciplinary field research. Are these kinds, different age forms; are they due to hardly perceptible soil or other ecological factors; are they the result of growing in semi-open or secondary situations, as opposed to the dense forest; are the specimens taken from various parts of the liana, are the cultivated specimens specially selected clones with varying chemical composition and, consequently, varying physiological effects; or are they chemovars?” (See Schultes, 1986)
Regional Origins of Plant-based Tryptamine + Beta-carboline Synergies and Migration Patterns for 3 Key Species of the Yagé Complex
Origins of the Modern Brew
Some of the writings by Schultes and other researchers, as well as herbarium collections from eastern Ecuador reference the Payaguaje family lineage (and related branches, see Payaguaje 1990 and 2007). They are amongst many notable aboriginal families of the region. When researching the roots of the ayahuasca tradition, all indications seem to converge upon the area between the Río Aguarico and the Río Putumayo as a major region of diversity. The Tukanoan cultural areas on the upper Río Negro in Colombia would be another epicenter of diversity. Additionally, researcher Gale Highpine, places the origin of ayahuasca (in the popular form known today) in the Northwest Amazon where the Río Napo meets the Amazon River.
Ayahuasca = Banisteriopsis caapi (Photos and Illustrations: N. Logan), 1 – Banisteriopsis caapi var. ourinho from Brazil, 2 – Banisteriopsis caapi var. cielo flowers, 3 – Banisteriopsis caapi immature fruit, 4 – Banisteriopsis caapi var. DaVine fresh flowers, 5 – Banisteriopsis caapi trunk bark, 6 – Banisteriopsis caapi var. tukunacá from Hawaii, 7 – Banisteriopsis caapi var. DaVine close-up of flower
Due to a host of factors that are political, ecological, and cultural, for the last several decades it has been relatively easy for western explorers to access Ecuador. Since the vines cared for by these Ecuadorian indigenous families (who have a long-standing, well-established relationship with these plants and have handed them down person to person over many generations), they can be seen as a kind of reference standard against which other vines can be compared. Consider the northwest Amazon basin nestled up against the eastern Andes as the epicenter with spokes radiating mostly to the east and south of that point. The logic here is that the discovery and refinement of the technology we know today as modern yagé or ayahuasca, was born in this epicenter and then slowly spread downriver and traversed the eastern slopes of the Andes. The implication is that most vines found further away from this region are derivations of the legacy vines and their hybrids originating at the epicenter. By cataloguing the attributes of legacy vines and their ethno-categories, a reference can be built for determining the pedigree of any vine of The Yagé Complex, a term that can be used to refer to the numerous named cultivated varieties of Banisteriopsis caapi and related Malpighiaceous vines incorporated in yagé/ayahuasca preparations with analogous effects. B. caapi members of The Yagé Complex can be categorized according to four fundamental morphological features of these related plants: they have either smooth or swollen nodes; they grow as either low bushes or high climbing vines. There are many hybrids that intergrade within these four features, sharing characteristics that result in new cultivars.
Tara = Banisteriopsis caapi var. tara (Photos: N. Logan), 1 – abaxial leaf glands flanking petiole at base of lamina, 2a – trunk bark, 2b – branch, 3a – fresh flower, 3b – mauve (#E0B0FF) flowers that fade to white, 4 – leaves with complex venation (twice divided)
The Payaguaje Collection of Legacy Vines
Flower color and structure are important macroscopic features for identifying cultivars of B. caapi. Ayahuasca typically has flowers with pink petals that fade to white or creamy yellow when old, yet great variation exists. From the Río Aguarico of Ecuador has come a very special collection of vines that serve as a reference for The Yagé Complex. E. Jean Langdon’s work describes the people from this region who identify as Siekopai and speak the Paicoca language. Linguistic evidence links three of these legacy vines (tara, tzinca and wai yagé) to this region of Ecuador in Sucumbíos Province. The tara vine (which means bone in Paicoca) has long straight stem growth, smooth nodes and a flower with mauve petals that fade to white. The name may be a reference to the long slender portion of a human bone (diaphysis). According to Jonathon Miller Weisberger (in an accompanying Microcosms article), when tara yagé is prepared, the stems are pounded until the outer bark is completely removed and what remains resembles the inner part of the bone. According to Weisberger, tara yagé is so powerful that it is both cultivated and prepared far away from the community following strict protocols. The tzinca vine has large swollen nodes and white flower petals with a pink splotch running through the middle. It is also known for producing strong physical effects in those who drink it such as shaking and purging. Tzinca in the Paicoca language refers to the bulbous ends of a human long bone (epiphysis) alluding to the large swollen nodes of this vine. Wai yagé grows as a mounded shrub with flowers, similar to “Tzinca”. Weisberger relates that the word “Wai” refers to meat, fish or game in general. This vine is prepared and ingested close to home, sometimes uncooked without additional admixtures (elaborated in a way similar to how kavakava is prepared in the Pacific islands). Wai yagé is consumed for the purpose of tracking the activities of animals. Branches and leaves of “yagé oco” are often added to one or more of the previously mentioned vines to produce “yagé”. The Payaguaje legacy vines, along with many related medicinal plants were collected with permission and encouraged to be spread and consumed. Various parts of the collection have made their way to the US mainland and Hawaii and are now grown across the southern US. The clearest message disseminated with this collection (across time and cultures) is, “keep drinking”. None of the plants in this collection are for sale, and each plant has been shared lovingly from the hands of one gardener to another the way it has been for millennia. It is in this spirit that confocal images of these plants from The Yagé Complex appear in this ecodigital repository that constitutes Microcosms: A Homage to Sacred Plants of the Americas.
Wai Yagé – Banisteriopsis caapi var. wai yagé (Photos: T. Baldwin and N. Logan), 1 – Leaf with abaxial glands around the margin, 2 – vine in habit, 3 – Flowers, 4 – stem cross section, 5 – swollen stems with short distance between nodesWai Yagé – Banisteriopsis caapi var. wai yagé, rare to see in flower, here pictured growing on the big Island of Hawaii, photo Neil Logan
Caapi by Any Other Name…
Most westerners understand ayahuasca as simply orally-activated dmt, with the vine merely serving to supply this function. Westerners tend to look for and expect certain effects from encounters with “psychedelics” (of which yagé is considered a member). For this reason, there is a focus on the dmt-containing plants. However, it’s important for western enthusiasts of ayahuasca to remember that, regardless which of the more than a hundred potential admixture plants are added to the brew, it’s still referred to by the name of the vine called ayahuasca or yagé. The name of the brew thus reflects its primary ingredient. Spruce’s notes demonstrate there were different recipes for the brew for different purposes and by different people. Documentation of those early encounters by westerners consistently report B. caapi as the principal ingredient.
Resolving the Enigma
The original materials that best exemplify a species concept are referred to as the type and lectotype. Brazilian botanists working on Banisteriopsis caapi and its ethnotaxonomic varieties have only recently resolved these fundamental elements of species taxonomy (See de Oliveira et al and de Souza et al). Furthermore, vines from many genera of the neotropical Malpighiaceae are known to be used by people for traditional medicine and/or known to contain bioactive compounds. Examples of these genera include: Alicia, Bronwenia, Hirae, Tetrapterys, Banisteriopsis, Diplopterys, Callaeum, Mezia, Heteropterys, Glicophyllum, Stygmaphyllon, and others. The neotropical Malpighiaceae have been in taxonomic revision since the 1980’s and are expected to continue being researched and assessed for some time to come.
Tzinca = Banisteriopsis caapi var. tzinca (Photos: T. Baldwin and N. Logan), 1 – leaf, 2 – vine in habit displaying closely spaced swollen nodes 3 – flowers, 4 – leaf gland, 5 – leaf gland on base of lamina
The difficulty of segregating these vines was, precisely, the enigma faced by Schultes. He was a botanist who collected and correctly identified thousands of plants in hyper-diverse tropical rainforests and yet these particular vines still confounded him. Botanists (Anderson and Gates) only recognize B. caapi as a single taxon with no botanically-valid varieties. Indigenous cultures and academics and preparers affiliated with the Santo Daime church in Brazil (see Monteles) have named varieties that number in the dozens. To segregate the two main varieties tucunacá (smooth nodes) and caupuri (swollen nodes) from one another, scientists have resorted to using light microscopes to peer into palisade parenchyma cells and vascular bundles. These botanists are only surveying morphological features such as flowers, leaves, bark, growth habit, microscopic cellular structure, etc., whereas the other groups (indigenous cultures and entheogenic churches in Brazil) have more experiential ways of knowing and categorizing that, in addition to morphology, can include sap color, number of lobes in the cross section of a stem, flavor, scent of leaves, physical and psycho-spiritual effects, amongst other characteristics. According to Schultes, their identity “depends on the conjunction of botanical features, chemical effects of the mode of preparation and cultural suggestions in the visions experienced.” Each cultivar is known to have their own unique effects and context for use, which are often intertwined with their respective rituals.
“Each plant has a spiritual guardian and a shaman owner, and shamans often trade kinds. Furthermore, if a shaman finds a wild liana in the forest, he will prepare a drink to ascertain its worth for inclusion in his own repertoire, especially in regard to what visions it can induce; if he takes a cutting, he will then and there name and classify it [….] Further exploration between this conjunction of botany-chemistry-culture warrants further investigation [….] It is still an enigma.” (Schultes 1986)
Yagé oco = Diplopterys longialata var. huambisa (Photos: T. Baldwin and N. Logan) 1 – cross section of petiole, 2 – large nectaries at the base of the lamina, 3 – vine in habit, 4 – multiple meristematic shoots per node, 5 – flowers, 6 – stem cross section, 7 – trunk with deep vertical inclusions, 8 – mature leaf and fruit
All-in-One Vine
It has long been hypothesized there may have been an original proto-vine that contained beta-carboline and tryptamine alkaloids all in a single, easy-to-consume-plant (something akin to Psilocybe mushrooms). During his search for caapi-pinima, Schultes consumed a cold water extraction of bark scrapings from Tetrapterys methystica and collected samples of the plant source. Much of his collection was lost but some was salvaged. The fragments that are left seem to indicate another epithet: Glicophyllum stylopterum (A.Juss.) R.F.Almeida. Little is known about this species from modern research. However, in Brazil, a closely- related species that is considered another kind of cabi, T. mucronata, recently has been investigated because Brazilian churches there are currently using it. The chemistry is such that it could be used as the sole ingredient in an ayahuasca-like preparation. The presences of 5Meo-DMT, 5Meo-n-methyl-typtamine, bufotenine, and 2-methyl-6-methoxy-1,2,3,4-tetrahydro-β-carboline have been detected in this species. It is considered to be something only for specialists because it is potentially very dangerous due to possible cardio-activity (Queiroz 2013).
Diplopterys cabrerana vs. D. longialata (Photographs by N. Logan with permission from the NYBG.) 1a – D. cabrerana fruit, 1b – D. cabrerana type collection: Schultes and Cabrera 17459 Rio Pira Paraná , Colombia, Sept. 10th 1952, 2a – D. longialata var. Huambisa fruit, 2b – D. longialata collected by Limbach, M.D., C.F., No. 148, Morona Santiago, Ecuador, March 1, 1990.
Diplopterys cabrerana Confusion
Another candidate for the all-in-one brew would seem to come from B. rusbyana (syn. D. longialata). This vine has long been conflated with Diplopterys cabrerana and appears to be the more commonly-employed admixture of the two in modern brews. There are herbarium collections of this species indicating that the trunk can be used to prepare a beverage (See Velarde-Núñez). Phytochemical analysis of this species found dmt in the leaves and bark as well as 2-methyl-tetrahydro-beta-carboline in the branches and trunk bark. Experience with this brew (where all parts of this vine are used) indicate the flavor is putrid and undrinkable, leading to the hypothesis that someone, somewhere along the way, figured out that the leaves of this vine (D. longialata) could be mixed with the bark that is relatively sweet-tasting (at least in comparison to that of B. rusbyana bark) of B. caapi, to produce a palatable tea. Might this explain how the modern yagé brew was born? This major technological advancement could have catalyzed the recent usage of caapi since a wide range of people could accept the flavor of this brew. Additionally, the National Cancer Institute (USA) funded a project in Ecuador led by C. F. Limbach in which two of their collections (“natem” B. caapi and “yaji” D. longialata) brewed together and reduced to a concentrated liquid extract is consumed to treat “various severe illness, incl. TB, malaria, yellow fever, and symptoms resembling female genitourinary carcinomas and basal cell carcinomas” (See Limbach, herbarium specimen, NYBG).
The strong morphological similarities between D. longialata and D. cabrerana and their ethnomedicinal uses have made proper collection and identification of the two very difficult. Their frequent usage keeps them in a near perpetual state of vegetative growth. For this reason, the vines rarely flower or reproduce, making their identification next to impossible.
Other vines to know part 1 (Photos: N. Logan), 1a&b – purgahuasca = Alicia anisopetala flowers and leaves, 2 – mii = Banisteriopsis muricata leaves, 3a,b,&c – Neidenzuella stannea – easily confused with desirable admixtures – flowers, trunk, leaves, 4 – huilca bejuco = Diplopterys lutea leaves
Other Vines to Know
The pressure due to overharvesting of traditional B. caapi is now so great that alternative (back up) sources are being sought. D. pubipetala has become popular in recent times as an analogue of B. caapi. Its bark is considered a functional source of harmine and the plant produces compounds in the leaves that are being studied for their ability to treat cancer. Alicia anisopetala known as purgahuasca is used (like “snake head soup” made from B. caapi leaves) as a pre-ceremony cleanse. There is also a powerful vine with blue flowers (species unknown) and the “white ayahuasca” aka “Yawarpanga” which appears to be from the genus Aristolochia – in a completely different family (Aristolochiaceae) from yagé. This vine is used as a ritual emetic before ceremonies that are focused on healing drug addiction. Then there is the look-alike vine Neidenzuella stannea with leaves that could be confused with Diplopterys cabrerana and D.longialata or with Banisteriopsis muricata with shiny two-toned (discolor) leaves. This vine contains MFA (mono floro-acetate), a non-volatile compound that doesn’t boil off during the brewing process and acts as a respiratory suppressant known to kill cows! To add to the mix, R.E. Schultes wrote, “The plant Hawkesiophyton ochraceum (Cuatrec.) A.Orejuela & C.I.Orozco (syn. Juanulloa ochracea) (Solanaceae), which contains the alkaloid parquina is called Ayahuasca in the Colombian Putumayo, and is added to Banisteriopsis drinks.” Finally, Mansoa alliaceae., (Bignoniaceae), known locally as ajosacha (wild garlic), is another vine commonly added to caapi brews. Its leaves smell like garlic and are sometimes mistaken for Diplopterys cabrerana leaves. The roots are used to cleanse the body of parasites and color the brew yellow.
Other vines to know Part 2 (Photos: N. Logan), 1 – ajo sacha = Mansoa alliacea leaves, 2 – white ayahuasca aka Yawarpanga = Aristolochia sp. leaves, 3 – caabi = Callaeum antifebrile, 4 – Diplopterys pubipetala leaves, 5 – ayahuasca = Hawkesiophyton ochraceum flowers
Supporting South American Indigenous Cultures
What is the meaning of these legacy vines outside of their cultural and ecological contexts? The situation in the Amazon is much more dire than is reflected in the media. Friends in the front-line trenches of reforestation in the Amazon and Cerrado regions of Peru and Brazil report there is only ~30% of the Amazon left intact, and only 20% of the Cerrado! Some organizations in Ecuador claim there could be complete deforestation there by 2025.The best line of defense against deforestation is to support the forest-based cultures whose identity and livelihoods are intimately intertwined with the ecosystem within which it is embedded.To his credit, rather than continuing to exact more pressure upon wild collected resources, L. Miller did plan to cultivate the vine to supply industry, which was potentially a good thing. However, patenting a culturally significant species (as his own) is certainly not appropriate. Moving forward will require shifting the current unsustainable approach to one that is more equitable and that duly honors the worldview of indigenous peoples. What’s at stake is more than any single species. With the loss of biological and cultural diversity, we collectively lose resiliency in the face of an ever-changing world. Indigenous experience points to these vines as cross-kingdom, communication technologies, instrumental to understanding and integrating with ecosystems. Tools of this kind will help us navigate the ecological catastrophe that humanity (collectively) has been creating over the past several hundred years.
Growing caapi (Photos: N. Logan), 1, 2 & 3 caapi var. ourinho growing on a rebar dome with chacruna beneath *note the leaf litter deposit, 4. Teepee of wooden poles for trellising caapi vines = safe and responsible (Illustration by N. Logan), 5. Sweet caapi growing on a native tree in Hawaii = dangerous irresponsible – will accelerate tree fall *if using trees for trellis, plant vines on the upwind side
Yagé is not only a miraculous plant teacher. The cultivation of B. caapi can also help regenerate forest ecosystems. Rather than allowing vines to grow haphazardly, wreaking havoc on the new ecosystems into which they are introduced, instead they can be trellised on domes with chacruna (Psychotria viridis) growing underneath. Planting vines in a system that is well managed has the potential to regenerate soil and bring back the forest. Using longstanding traditions of agroforestry, together, we can develop regenerative agroecosystems that produce raw materials to feed our supply chains. This will slow the need to further deplete intact forest ecosystems and empower indigenous people to manage their natural resource base in a manner that honors their heritage.
Tara = Banisteriopsis caapi var. tara (Photos: N. Logan), 1 – abaxial leaf glands flanking petiole at base of lamina, 2a – trunk bark, 2b – branch, 3a – fresh flower, 3b – mauve (#E0B0FF) flowers that fade to white, 4 – leaves with complex venation (twice divided)
Conclusion
Vines of The Yagé Complex play an important ecological and cultural role in the lives of people from the Amazon basin of South America. Shaped by the forces of continental drift, subsequent mountain building and human selection, these vines contain great diversity of form and function – a caapi for every occasion. As these lianas spread out across the continent from their point of origin, many new hybrids have been created that complicate identifying the pedigree and therefore application of unidentified vines. The morphological similarities between related taxa have perplexed even expert botanists. In recent decades, that confusion has been exploited, resulting in a patent awarded to a non-indigenous person with no acknowledgement given to the people responsible for introducing the west to these organisms in the first place. Over-exploitation of wild natural resources and deforestation have become undeniably serious threats to ecosystems and cultures who have coexisted and thrived together in the Amazon for millennia. Western culture must rectify its relationship with indigenous cultures on the basis of equality and respect in order to then integrate the wisdom of yagé to avoid global ecological catastrophe.
Photos
Diplopterys cabrerana “Yagé ocó” photo by Jonathon S. Miller WeisbergerDiplopterys cabrerana “Yagé ocó” young leaves, by Jonathon S. Miller WeisbergerDiplopterys longialata var Huambisa “Yagí” by Jonathon S. Miller WeisbergerDiplopterys longialata var Huambisa “Yagí” young leaves and extra floral nectaries, photo by Neil Logan
Bibliography
Davis, Charles C., Peter W. Fritsch, Charles D. Bell, and Sarah Mathews. “High Latitude Tertiary Migrations of an Exclusively Tropical Clade: Evidence from Malpighiaceae,” International Journal of Plant Sciences (Tropical Intercontinental Disjunctions) 165.S4 (July 2004): S107-S121.
Davis, Charles C. and William R. Anderson. “A Complete Generic Phylogeny of Malpighiaceae Inferred from Nucleotide Sequence Data and Morphology.” American Journal of Botany 97.12 (December 2010): 2031-2048. https://doi.org/10.3732/ajb.1000146
de Oliveira, R.C., Sonsin-Oliveira, J., dos Santos, T.A.C., Simas e Silva, M., Fagg, C.W. and Sebastiani, R. (2021), “Lectotypification of Banisteriopsis caapi and B. quitensis (Malpighiaceae), names associated with an important ingredient of Ayahuasca,” Taxon, 70: 185-188. https://doi.org/10.1002/tax.12407
de Oliveira, R.C., Behrens, C.S.B., Nagamine-Pinheiro, N. et al. “Ethnobotany and Wood Anatomy of Banisteriopsis caapi Ethnotaxa and Diplopterys cf. pubipetala, Components of Ayahuasca in Brazilian Rituals.” Economic Botany77 (2023): 18–47. https://doi.org/10.1007/s12231-023-09567-w
de Souza, Raniely Miranda et al. “Lectotypification and neotypification of names related toBanisteriopsis caapi(Malpighiaceae): a contribution to understanding of Ayahuasca,” Phytotaxa 585.1 (2023).
Díaz León, Germán Leonardo. “NUESTRO YAJÉ… ¿TU DA VINE?: Elementos económicos, culturales y legales del debate alrededor de la propiedad intelectual sobre los Saberes Tradicionales”. Trabajo de Grado para Optar al Título de Sociólogo. Departamento de Sociología, Facultad de Ciencias Humanas, Universidad Nacional de Colombia. 20 de enero de 2006.
Gates, Bronwen. “Banisteriopsis, Diplopterys (Malpighiaceae)”. Flora Neotropica 30 (Feb. 18, 1982): 1-237. (Published by New York Botanical Garden Press on behalf of Organization for Flora Neotropica). Stable URL: https://www.jstor.org/stable/4393754
Langdon, E. Jean. “Las clasificaciones del yagé dentro del grupo Siona: Etnobotánica, etnoquímica e história.” América Indígena 46 (1986): 101–116.
Limbach, M.D., C.F., Diplopterys longialata, Province of Morona Santiago, Medicinal Plants of the Shuar, No. 148 and CFL139, March 1, 1990. (Herbarium specimen at the NYBG)
Luz, Thalita Zanquetta, Antonio Saulo Cunha-Machado and Jacqueline da Silva Batista. “First DNA barcode efficiency assessment for an important ingredient in the Amazonian ayahuasca tea: mariri/jagube, Banisteriopsis (Malpighiaceae)”
Genet Resour Crop Evol (20 December 2022): https://doi. org/10.1007/s10722-022-01522-3
Miller Weisberger, Jonathon. “Rainforest Medicine, Preserving Indigenous Science and Biodiversity in the Upper Amazon” Berkeley California, North Atlantic Books, 2013
Monteles, Ricardo. “Eu venho da floresta”: A sustentabilidade das plantas sagradas amazônicas do Santo Daime. Universidade Federal do Amazonas (Manaus) (fevereiro 2020). Tese (Doutorado em Ciências do Ambiente e Sustentabilidade na Amazônia). https://tede.ufam.edu.br/bitstream/tede/7682/2/Tese_RicardoMonteles_PPGCASA.pdf
Nagamine-Pinheiro, Nívea et al. “Vegetative anatomy, morphology and histochemistry of three species of Malpighiaceae used in analogues of the Amazonian psychoactive beverage Ayahuasca,” Flora 275 (2021): https://doi.org/10.1016/j.flora.2020.151760
Payaguaje, Fernando. El bebedor de yagé. Shushufindi—Río Aguarico, Ecuador: Vicariato Apostólico de Aguarico, 1990.
Press, Sara V. “Ayahuasca on Trial: Biocolonialism, Biopiracy, and the Commodification of the Sacred.” History of Pharmacy and Pharmaceuticals 63.2 (January 2022): 328-353.
Queiroz. Marcos Marçal Ferreira “ldentificaçào dos inibidores de acetilcolinesterase em Tetrapterys mucronata Cav. (Malpighiaceae) e comparaçào quali e quantitativa dos derivados triptaminicos presentes na espécie eni estudo e Ayahuasca” Araraquara: [s.n], 2013.
Queiroz, M. M. F. et al. “Chemical Composition of the Bark of Tetrapterys mucronata and Identification of Acetylcholinesterase Inhibitory Constituents,” Journal of Natural Products 77.3 (2014): 650-656. https://doi.org/10.1021/np401003p
Queiroz, M. M. F. et al. “LC-MS/MS Quantitative Determination of Tetrapterys mucronata Alkaloids, A Plant Occasionally Used in Ayahuasca Preparation,” Phytochemical Analysis 26.3 (2015): 183-188. DOI: 10.1002/pca.2548
Schultes, Richard Evans. “Recognition of Variability in Wild Plants by Indians of the Northwest Amazon: An Enigma,” Journal of Ethnobiology 6.2 (Winter 1986): 229-238.
Spruce, Richard. “On Some Remarkable Narcotics of the Amazon Valley and Orinoco.” Ocean Highways: The Geographical Review 55.1 (1873): 184-193.
Torres, Constantino Manuel. “From Beer to Tobacco: A Probable Prehistory of Ayahuasca and Yagé,” in Dennis J. McKenna, ed. Ethnopharmacologic Search for Psychoactive Drugs: II. Santa Fe, New Mexico: Synergetic Press, 2018: 36-54.
Velarde Núñez, O. Banisteriopsis rusbyana Peru: Department of San Martin, Tarapoto Province, Tarapoto, alt. 600m. Fickle climber. Cultivated. Its trunk is used to prepare a hallucinogenic and purgative drink. N.v. “ayahuasca amarilla” 8/2/1958 O. Velarde Núñez 6589 (Herb. Morris Arb.)
Vickers, William T. Los sionas y secoyas: su adaptación al ambiente amazónico. Quito: Abya-Yala, 1989.
Vickers, William T. and Timothy Plowman. “Useful Plants of the Siona and Secoya Indians of Eastern Ecuador.” Fieldiana Botany (N.S.) 15 (1984):1–63.
Werneck, Beatriz et al. “Biodiversity of β-Carboline Profile of Banisteriopsis caapi and Ayahuasca, a Plant and a Brew with Neuropharmacological Potential,” MDPI (Plants) 9. 870 (9 July 2020): doi:10.3390/plants9070870
Wiser, Glenn. “U.S. Patent and Trademark Office Reinstates Ayahuasca Patent: Flawed Decision Declares Open Season on Resources of Indigenous Peoples,” (June 25, 2001). (Author is Staff Attorney, Center for International Environmental Law).
About the Author
Neil Logan is the Innovations Officer for AgroforestryX and co-creator of the AgroforestryX design tool. AgroforestryX assists farmers, land managers, and conservation project planners to assess, design and manage dynamic multistory agroforestry systems. He is personally passionate about agro-successional restoration systems that produce medicinal, timber, and food crops to offset the costs of regenerating ecosystems. In addition to his work on Malpighiaceae, he has done extensive research on Prosopis, a summary of which can be read in the Microcosms Plant Index.